

 journal menu
journal menu organic compounds
organic compounds





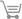
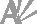


Supporting information
 | Crystallographic Information File (CIF) https://doi.org/10.1107/S010827010100261X/vj1132sup1.cif |
 | Structure factor file (CIF format) https://doi.org/10.1107/S010827010100261X/vj1132Isup2.hkl |
CCDC reference: 164668
Colourless single crystals of the amino acid were grown as fine transparent needles from a saturated aqueous solution. The density was determined by the flotation method using a liquid mixture of carbon tetrachloride and xylene.
All the H atoms were generated geometrically and treated with a riding model with Uiso fixed at 1.2Ueq of the bonded atoms or 1.5Ueq for amino and methyl groups.
Data collection: CAD-4 Software (Enraf-Nonius, 1989); cell refinement: CAD-4 Software; data reduction: CAD-4 Software; program(s) used to solve structure: SHELXS97 (Sheldrick, 1990); program(s) used to refine structure: SHELXL97 (Sheldrick, 1997); molecular graphics: PLATON (Spek, 1999); software used to prepare material for publication: SHELXL97.
![[Figure 1]](https://journals.iucr.org/c/issues/2001/05/00/vj1132/vj1132fig1thm.gif)
| Fig. 1. The molecular structure, with atom-numbering scheme and 50% probability displacement ellipsoids. |
![[Figure 2]](https://journals.iucr.org/c/issues/2001/05/00/vj1132/vj1132fig2thm.gif)
| Fig. 2. Packing diagram of the molecule viewed down the b axis. |
| C3H7NO2 | Dx = 1.399 Mg m−3 Dm = 1.39 Mg m−3 Dm measured by floatation |
| Mr = 89.10 | Cu Kα radiation, λ = 1.54180 Å |
| Orthorhombic, Pna21 | Cell parameters from 25 reflections |
| a = 12.0263 (17) Å | θ = 15–27° |
| b = 6.0321 (9) Å | µ = 1.00 mm−1 |
| c = 5.829 (2) Å | T = 293 K |
| V = 422.88 (19) Å3 | Fine needles, colourless |
| Z = 4 | 0.42 × 0.24 × 0.18 mm |
| F(000) = 192 |
| Enraf-Nonius CAD-4 diffractometer | 417 reflections with I > 2σ(I) |
| Radiation source: fine-focus sealed tube | Rint = 0.000 |
| Graphite monochromator | θmax = 67.6°, θmin = 7.4° |
| ω–2θ scans | h = −14→0 |
| Absorption correction: ψ scan (North et al., 1968) | k = 0→7 |
| Tmin = 0.97, Tmax = 0.99 | l = 0→7 |
| 422 measured reflections | 2 standard reflections every 60 min |
| 422 independent reflections | intensity decay: 2% |
| Refinement on F2 | Hydrogen site location: inferred from neighbouring sites |
| Least-squares matrix: full | H-atom parameters constrained |
| R[F2 > 2σ(F2)] = 0.025 | w = 1/[σ2(Fo2) + (0.0512P)2 + 0.0571P] where P = (Fo2 + 2Fc2)/3 |
| wR(F2) = 0.072 | (Δ/σ)max < 0.001 |
| S = 1.09 | Δρmax = 0.14 e Å−3 |
| 422 reflections | Δρmin = −0.15 e Å−3 |
| 56 parameters | Extinction correction: SHELXL97 (Sheldrick, 1997), Fc*=kFc[1+0.001xFc2λ3/sin(2θ)]-1/4 |
| 1 restraint | Extinction coefficient: 0.012 (3) |
| Primary atom site location: structure-invariant direct methods | Absolute structure: Flack H D (1983), Acta Cryst. A39, 876-881 |
| Secondary atom site location: difference Fourier map |
| C3H7NO2 | V = 422.88 (19) Å3 |
| Mr = 89.10 | Z = 4 |
| Orthorhombic, Pna21 | Cu Kα radiation |
| a = 12.0263 (17) Å | µ = 1.00 mm−1 |
| b = 6.0321 (9) Å | T = 293 K |
| c = 5.829 (2) Å | 0.42 × 0.24 × 0.18 mm |
| Enraf-Nonius CAD-4 diffractometer | 417 reflections with I > 2σ(I) |
| Absorption correction: ψ scan (North et al., 1968) | Rint = 0.000 |
| Tmin = 0.97, Tmax = 0.99 | 2 standard reflections every 60 min |
| 422 measured reflections | intensity decay: 2% |
| 422 independent reflections |
| R[F2 > 2σ(F2)] = 0.025 | 1 restraint |
| wR(F2) = 0.072 | H-atom parameters constrained |
| S = 1.09 | Δρmax = 0.14 e Å−3 |
| 422 reflections | Δρmin = −0.15 e Å−3 |
| 56 parameters | Absolute structure: Flack H D (1983), Acta Cryst. A39, 876-881 |
Geometry. All e.s.d.'s (except the e.s.d. in the dihedral angle between two l.s. planes) are estimated using the full covariance matrix. The cell e.s.d.'s are taken into account individually in the estimation of e.s.d.'s in distances, angles and torsion angles; correlations between e.s.d.'s in cell parameters are only used when they are defined by crystal symmetry. An approximate (isotropic) treatment of cell e.s.d.'s is used for estimating e.s.d.'s involving l.s. planes. |
Refinement. Refinement of F2 against ALL reflections. The weighted R-factor wR and goodness of fit S are based on F2, conventional R-factors R are based on F, with F set to zero for negative F2. The threshold expression of F2 > σ(F2) is used only for calculating R-factors(gt) etc. and is not relevant to the choice of reflections for refinement. R-factors based on F2 are statistically about twice as large as those based on F, and R- factors based on ALL data will be even larger. |
| x | y | z | Uiso*/Ueq | ||
| O1 | 0.58960 (11) | 0.4862 (2) | 0.6045 (3) | 0.0356 (4) | |
| O2 | 0.68614 (12) | 0.2009 (2) | 0.7409 (3) | 0.0328 (4) | |
| C2 | 0.66363 (15) | 0.2231 (3) | 0.3380 (3) | 0.0224 (4) | |
| H2 | 0.7419 | 0.1804 | 0.3237 | 0.027* | |
| C1 | 0.64429 (13) | 0.3133 (3) | 0.5818 (3) | 0.0222 (4) | |
| C3 | 0.59233 (16) | 0.0216 (3) | 0.2916 (4) | 0.0347 (5) | |
| H3A | 0.6060 | −0.0308 | 0.1387 | 0.052* | |
| H3B | 0.5153 | 0.0605 | 0.3069 | 0.052* | |
| H3C | 0.6104 | −0.0930 | 0.3997 | 0.052* | |
| N1 | 0.63948 (12) | 0.3976 (2) | 0.1659 (3) | 0.0258 (4) | |
| H1A | 0.6815 | 0.5160 | 0.1942 | 0.039* | |
| H1B | 0.5680 | 0.4351 | 0.1741 | 0.039* | |
| H1C | 0.6544 | 0.3463 | 0.0261 | 0.039* |
| U11 | U22 | U33 | U12 | U13 | U23 | |
| O1 | 0.0407 (7) | 0.0418 (7) | 0.0242 (9) | 0.0146 (5) | 0.0024 (7) | −0.0047 (6) |
| O2 | 0.0415 (7) | 0.0416 (8) | 0.0152 (7) | 0.0080 (6) | −0.0017 (6) | 0.0006 (6) |
| C2 | 0.0256 (7) | 0.0287 (9) | 0.0130 (9) | 0.0029 (7) | 0.0010 (7) | −0.0009 (8) |
| C1 | 0.0210 (8) | 0.0319 (8) | 0.0136 (9) | −0.0013 (6) | 0.0004 (6) | −0.0026 (8) |
| C3 | 0.0457 (11) | 0.0340 (9) | 0.0245 (13) | −0.0057 (8) | −0.0022 (9) | −0.0036 (9) |
| N1 | 0.0300 (7) | 0.0326 (8) | 0.0148 (8) | 0.0002 (6) | 0.0003 (6) | 0.0002 (6) |
| O1—C1 | 1.240 (2) | C3—H3A | 0.9600 |
| O2—C1 | 1.254 (3) | C3—H3B | 0.9600 |
| C2—N1 | 1.483 (3) | C3—H3C | 0.9600 |
| C2—C3 | 1.512 (3) | N1—H1A | 0.8900 |
| C2—C1 | 1.540 (3) | N1—H1B | 0.8900 |
| C2—H2 | 0.9800 | N1—H1C | 0.8900 |
| N1—C2—C3 | 109.80 (17) | H3A—C3—H3B | 109.5 |
| N1—C2—C1 | 110.13 (15) | C2—C3—H3C | 109.5 |
| C3—C2—C1 | 111.30 (18) | H3A—C3—H3C | 109.5 |
| N1—C2—H2 | 108.5 | H3B—C3—H3C | 109.5 |
| C3—C2—H2 | 108.5 | C2—N1—H1A | 109.5 |
| C1—C2—H2 | 108.5 | C2—N1—H1B | 109.5 |
| O1—C1—O2 | 126.08 (19) | H1A—N1—H1B | 109.5 |
| O1—C1—C2 | 118.42 (19) | C2—N1—H1C | 109.5 |
| O2—C1—C2 | 115.50 (15) | H1A—N1—H1C | 109.5 |
| C2—C3—H3A | 109.5 | H1B—N1—H1C | 109.5 |
| C2—C3—H3B | 109.5 | ||
| N1—C2—C1—O1 | 16.3 (2) | N1—C2—C1—O2 | −163.97 (16) |
| C3—C2—C1—O1 | −105.69 (18) | C3—C2—C1—O2 | 74.0 (2) |
| D—H···A | D—H | H···A | D···A | D—H···A |
| N1—H1A···O2i | 0.89 | 1.96 | 2.817 (2) | 160 |
| N1—H1B···O1ii | 0.89 | 2.00 | 2.865 (2) | 165 |
| N1—H1C···O2iii | 0.89 | 1.92 | 2.804 (3) | 173 |
| C2—H2···O1iv | 0.98 | 2.67 | 3.566 (3) | 153 |
| Symmetry codes: (i) −x+3/2, y+1/2, z−1/2; (ii) −x+1, −y+1, z−1/2; (iii) x, y, z−1; (iv) −x+3/2, y−1/2, z−1/2. |
Experimental details
| Crystal data | |
| Chemical formula | C3H7NO2 |
| Mr | 89.10 |
| Crystal system, space group | Orthorhombic, Pna21 |
| Temperature (K) | 293 |
| a, b, c (Å) | 12.0263 (17), 6.0321 (9), 5.829 (2) |
| V (Å3) | 422.88 (19) |
| Z | 4 |
| Radiation type | Cu Kα |
| µ (mm−1) | 1.00 |
| Crystal size (mm) | 0.42 × 0.24 × 0.18 |
| Data collection | |
| Diffractometer | Enraf-Nonius CAD-4 diffractometer |
| Absorption correction | ψ scan (North et al., 1968) |
| Tmin, Tmax | 0.97, 0.99 |
| No. of measured, independent and observed [I > 2σ(I)] reflections | 422, 422, 417 |
| Rint | 0.000 |
| (sin θ/λ)max (Å−1) | 0.600 |
| Refinement | |
| R[F2 > 2σ(F2)], wR(F2), S | 0.025, 0.072, 1.09 |
| No. of reflections | 422 |
| No. of parameters | 56 |
| No. of restraints | 1 |
| H-atom treatment | H-atom parameters constrained |
| Δρmax, Δρmin (e Å−3) | 0.14, −0.15 |
| Absolute structure | Flack H D (1983), Acta Cryst. A39, 876-881 |
Computer programs: CAD-4 Software (Enraf-Nonius, 1989), CAD-4 Software, SHELXS97 (Sheldrick, 1990), SHELXL97 (Sheldrick, 1997), PLATON (Spek, 1999), SHELXL97.
| D—H···A | D—H | H···A | D···A | D—H···A |
| N1—H1A···O2i | 0.89 | 1.96 | 2.817 (2) | 160 |
| N1—H1B···O1ii | 0.89 | 2.00 | 2.865 (2) | 165 |
| N1—H1C···O2iii | 0.89 | 1.92 | 2.804 (3) | 173 |
| C2—H2···O1iv | 0.98 | 2.67 | 3.566 (3) | 153 |
| Symmetry codes: (i) −x+3/2, y+1/2, z−1/2; (ii) −x+1, −y+1, z−1/2; (iii) x, y, z−1; (iv) −x+3/2, y−1/2, z−1/2. |




DL-Alanine, (I), is one of the few amino acids for which accurate X-ray crystal structure is not known. Previous work on this amino acid reports the cell dimensions (Bernal, 1931) and X-ray crystal structures derived from two-dimensional intensity data (Levy & Corey, 1941; Donohue, 1950). We report here an accurate determination of the crystal structure of DL-alanine at room temperature. This structure represents a rare case of an amino acid racemate crystallizing in a non-centrosymmetric space group. Another such structure is DL-tyrosine (Mostad & Romming, 1973). \sch
The DL-alanine molecule exists as a zwitterion. The C—O distances in the caboxylate group are unequal presumably due to the participation of one of the O atoms (O1) in one hydrogen bond and the other (O2) in two. The C—N distance, formerly thought to be unusually shorter by Levy & Corey (1941) with a value of 1.427 Å, is found to be 1.483 (3) Å in the present work. This is slightly less than the value of 1.496 Å quoted by Donohue (1950). The N atom deviates by 0.392 (5) Å from the carboxylate plane and the methyl carbon deviates by 1.356 (4) Å, in the opposite direction.
The crystal structure is stabilized by a network of characteristic head-to-tail hydrogen-bond sequences. The structure contains three types of such sequences viz. S2 (straight sequence along the c axis with O2 of the carboxylate group as acceptor), Z1 (zigzag sequence along the 21 screw axis with O1 of the carboxylate group as acceptor) and DL2 (zigzag-DL sequence among the glide related molecules with O2 of the carboxylate group as acceptor) (Suresh & Vijayan, 1983). While the sequences S2 and Z1 connect molecules in each layer, the zigzag-DL sequence connect alternating layers, each containing one isomer. The direction of the DL2 sequence is parallel to the plane of the amino acid layers. There is a striking similarity between this structure and that of its L-isomer (Simpson & Marsh, 1966; Destro et al., 1988) which is not uncommon in most other hydrophobic amino acids too (Soman & Vijayan, 1989). The cell dimensions of L– and DL– isomers are nearly identical. Both structures belong to orthorhombic system, but the space group is P212121 for the L-isomer and Pna21 for the racemate. Further, the arrangements of molecules within layers in the crystal structures of both L– and DL-alanine are identical. However, the DL2 sequence observed in the racemate is replaced by a Z2 sequence in its L-isomer. In addition, a weak C—H···O hydrogen bond, with the carboxylate oxygen O1 as acceptor, is observed among the glide-related molecules, interconnecting alternate layers, each containing one isomer.