

 journal menu
journal menu organic compounds
organic compounds







![[lambda]](/logos/entities/lambda_rmgif.gif) 5-furo[3,2-d][1,3,25]dioxaphosphinin-2-olate], C6H10N3O2+·C10H10N4O7P-, the Hoogsteen edge of the hypoxanthine (Hyp) base of cIMP and the Hyp face are engaged in specific amino acid-nucleotide (His
5-furo[3,2-d][1,3,25]dioxaphosphinin-2-olate], C6H10N3O2+·C10H10N4O7P-, the Hoogsteen edge of the hypoxanthine (Hyp) base of cIMP and the Hyp face are engaged in specific amino acid-nucleotide (His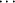 cIMP) recognition, i.e. by abutting edge-to-edge and by
cIMP) recognition, i.e. by abutting edge-to-edge and by ![[pi]](/logos/entities/pi_rmgif.gif) - stacking, respectively. The Watson-Crick edge of Hyp and the cIMP phosphate group play a role in nonspecific HiscIMP contacts. The interactions between the cIMP anions (anti/C3'-endo/trans-gauche/chair conformers) are realized mainly between riboses and phosphate groups. The results for this L-His-cIMP complex, compared with those for the previously reported solvated L-His-IMP crystal structure, indicate a different nature of amino acid-nucleotide recognition and interactions upon the 3':5'-cyclization of the nucleotide phosphate group.
- stacking, respectively. The Watson-Crick edge of Hyp and the cIMP phosphate group play a role in nonspecific HiscIMP contacts. The interactions between the cIMP anions (anti/C3'-endo/trans-gauche/chair conformers) are realized mainly between riboses and phosphate groups. The results for this L-His-cIMP complex, compared with those for the previously reported solvated L-His-IMP crystal structure, indicate a different nature of amino acid-nucleotide recognition and interactions upon the 3':5'-cyclization of the nucleotide phosphate group.Supporting information
 | Crystallographic Information File (CIF) https://doi.org/10.1107/S0108270112030041/qs3016sup1.cif |
 | Structure factor file (CIF format) https://doi.org/10.1107/S0108270112030041/qs3016Isup2.hkl |
CCDC reference: 899073
An MeOH–water solution (1:1 v/v) containing a 1:1 molar ratio of cyclic inosine 5'-phosphate sodium salt, Na(cIMP) (10 mg; Sigma) and L-histidine (L-His, 4.4 mg; ROTH) was evaporated slowly at room temperature for several days to give colourless columns of (I).
All H atoms were found in difference Fourier maps. In the final refinement cycles, they were positioned geometrically and treated as riding atoms, with C—H = 0.95–1.00 Å, N—H = 0.88–0.91 Å and O—H = 0.84 Å, and with Uiso(H) = 1.2Ueq(C,Nsp2) or 1.5Ueq(O,Nsp3).
Data collection: CrysAlis CCD (Oxford Diffraction, 2009); cell refinement: CrysAlis RED (Oxford Diffraction, 2009); data reduction: CrysAlis RED (Oxford Diffraction, 2009); program(s) used to solve structure: SHELXS97 (Sheldrick, 2008); program(s) used to refine structure: SHELXL97 (Sheldrick, 2008); molecular graphics: DIAMOND (Brandenburg, 2005); software used to prepare material for publication: SHELXL97 (Sheldrick, 2008).
![[Figure 1]](http://journals.iucr.org/c/issues/2012/08/00/qs3016/qs3016fig1thm.gif)
| Fig. 1. Views of the His cation and cIMP anion of (I), showing the atom-numbering schemes and the symmetry-independent N—H···O and C—H···N hydrogen bonds (dashed lines). Displacement ellipsoids are drawn at the 50% probability level. |
![[Figure 2]](http://journals.iucr.org/c/issues/2012/08/00/qs3016/qs3016fig2thm.gif)
| Fig. 2. Comparison of the conformations of (a) cIMP/IMP anions and (b) His cations in L-His–cIMP (black) and L-His–IMP.MeOH.H2O (Ślepokura & Petrus, 2010; light orange in the electronic version of the journal). The common reference points are the labelled atoms. In IMP, the relative orientation of the hypoxanthine (Hyp) base and ester atom O5' is stabilized by an intramolecular C8—H8···O5' hydrogen bond (dashed line). |
![[Figure 3]](http://journals.iucr.org/c/issues/2012/08/00/qs3016/qs3016fig3thm.gif)
| Fig. 3. (a The ribbon of His cations running down the a axis (b) and its specific His···cIMP edge-to-edge hydrogen bonds (thick red dashed lines) and π–π stacking (thin red dashed lines) with the cIMP ribbon. Intermolecular His···His and cIMP···cIMP interactions are shown as blue dashed lines. (Colours refer to the electronic version of the journal.) H atoms not involved in hydrogen bonds have been omitted for clarity. [Symmetry codes: (iv) x + 1, y, z; (v) x + 1/2, -y + 3/2, -z + 1.] |
![[Figure 4]](http://journals.iucr.org/c/issues/2012/08/00/qs3016/qs3016fig4thm.gif)
| Fig. 4. The complete set of His···cIMP interactions in (I). Specific and nonspecific hydrogen bonds are shown as heavy dashed lines and other contacts are shown as thin dashed lines. [Rephrasing OK?] H atoms not involved in hydrogen bonds have been omitted for clarity. [Symmetry codes: (ii) x - 1/2, -y + 1/2, -z + 1; (iii) -x + 3/2, -y + 1, z - 1/2; (vi) -x + 2, y + 1/2, -z + 3/2; (viii) x + 1/2, -y + 1/2, -z + 1.] |
| C6H10N3O2+·C10H10N4O7P− | F(000) = 1008 |
| Mr = 485.36 | Dx = 1.674 Mg m−3 |
| Orthorhombic, P212121 | Mo Kα radiation, λ = 0.71073 Å |
| Hall symbol: P 2ac 2ab | Cell parameters from 14525 reflections |
| a = 5.709 (2) Å | θ = 2.7–38.5° |
| b = 10.284 (3) Å | µ = 0.22 mm−1 |
| c = 32.795 (9) Å | T = 90 K |
| V = 1925.4 (10) Å3 | Column, colourless |
| Z = 4 | 0.12 × 0.07 × 0.04 mm |
| Oxford Xcalibur PX κ-geometry diffractometer with an Onyx CCD camera | 5114 independent reflections |
| Radiation source: Enhance (Mo) X-ray Source | 4778 reflections with I > 2σ(I) |
| Graphite monochromator | Rint = 0.032 |
| ω and ϕ scans | θmax = 30.0°, θmin = 2.7° |
| Absorption correction: multi-scan (CrysAlis RED; Oxford Diffraction, 2009) | h = −8→4 |
| Tmin = 0.949, Tmax = 1.000 | k = −12→14 |
| 17935 measured reflections | l = −46→46 |
| Refinement on F2 | Secondary atom site location: difference Fourier map |
| Least-squares matrix: full | Hydrogen site location: difference Fourier map |
| R[F2 > 2σ(F2)] = 0.030 | H-atom parameters constrained |
| wR(F2) = 0.077 | w = 1/[σ2(Fo2) + (0.0495P)2 + 0.2437P] where P = (Fo2 + 2Fc2)/3 |
| S = 1.04 | (Δ/σ)max = 0.001 |
| 5114 reflections | Δρmax = 0.35 e Å−3 |
| 300 parameters | Δρmin = −0.27 e Å−3 |
| 0 restraints | Absolute structure: from known absolute configuration and anomalous dispersion effects (Flack, 1983), with 1877 Friedel pairs |
| Primary atom site location: structure-invariant direct methods | Absolute structure parameter: 0.20 (6) |
| C6H10N3O2+·C10H10N4O7P− | V = 1925.4 (10) Å3 |
| Mr = 485.36 | Z = 4 |
| Orthorhombic, P212121 | Mo Kα radiation |
| a = 5.709 (2) Å | µ = 0.22 mm−1 |
| b = 10.284 (3) Å | T = 90 K |
| c = 32.795 (9) Å | 0.12 × 0.07 × 0.04 mm |
| Oxford Xcalibur PX κ-geometry diffractometer with an Onyx CCD camera | 5114 independent reflections |
| Absorption correction: multi-scan (CrysAlis RED; Oxford Diffraction, 2009) | 4778 reflections with I > 2σ(I) |
| Tmin = 0.949, Tmax = 1.000 | Rint = 0.032 |
| 17935 measured reflections |
| R[F2 > 2σ(F2)] = 0.030 | H-atom parameters constrained |
| wR(F2) = 0.077 | Δρmax = 0.35 e Å−3 |
| S = 1.04 | Δρmin = −0.27 e Å−3 |
| 5114 reflections | Absolute structure: from known absolute configuration and anomalous dispersion effects (Flack, 1983), with 1877 Friedel pairs |
| 300 parameters | Absolute structure parameter: 0.20 (6) |
| 0 restraints |
Geometry. All e.s.d.'s (except the e.s.d. in the dihedral angle between two l.s. planes) are estimated using the full covariance matrix. The cell e.s.d.'s are taken into account individually in the estimation of e.s.d.'s in distances, angles and torsion angles; correlations between e.s.d.'s in cell parameters are only used when they are defined by crystal symmetry. An approximate (isotropic) treatment of cell e.s.d.'s is used for estimating e.s.d.'s involving l.s. planes. |
Refinement. Refinement of F2 against ALL reflections. The weighted R-factor wR and goodness of fit S are based on F2, conventional R-factors R are based on F, with F set to zero for negative F2. The threshold expression of F2 > σ(F2) is used only for calculating R-factors(gt) etc. and is not relevant to the choice of reflections for refinement. R-factors based on F2 are statistically about twice as large as those based on F, and R-factors based on ALL data will be even larger. |
| x | y | z | Uiso*/Ueq | ||
| P1 | 0.38224 (6) | 0.35408 (4) | 0.353801 (10) | 0.01193 (8) | |
| O3 | 0.61881 (17) | 0.31050 (12) | 0.33964 (3) | 0.0150 (2) | |
| O5 | 0.30461 (19) | 0.31239 (12) | 0.39478 (3) | 0.0183 (2) | |
| O2' | −0.20278 (17) | 0.35658 (12) | 0.26364 (3) | 0.0142 (2) | |
| H2' | −0.2159 | 0.3305 | 0.2878 | 0.021* | |
| O3' | 0.18721 (17) | 0.30460 (11) | 0.32118 (3) | 0.0116 (2) | |
| O4' | 0.16333 (18) | 0.55904 (10) | 0.24623 (3) | 0.0130 (2) | |
| O5' | 0.36371 (19) | 0.50970 (11) | 0.35130 (3) | 0.0160 (2) | |
| O6 | 0.65016 (17) | 0.29662 (11) | 0.07526 (3) | 0.0140 (2) | |
| N1 | 0.3036 (2) | 0.20214 (13) | 0.09418 (3) | 0.0120 (2) | |
| H1 | 0.3073 | 0.1541 | 0.0720 | 0.014* | |
| N3 | 0.0814 (2) | 0.25197 (13) | 0.15336 (3) | 0.0129 (2) | |
| N7 | 0.5859 (2) | 0.46172 (13) | 0.15336 (3) | 0.0126 (2) | |
| N9 | 0.26574 (19) | 0.42739 (13) | 0.19164 (3) | 0.0105 (2) | |
| C1' | 0.0808 (2) | 0.45442 (15) | 0.22173 (4) | 0.0109 (3) | |
| H1' | −0.0663 | 0.4808 | 0.2074 | 0.013* | |
| C2' | 0.0302 (2) | 0.33863 (15) | 0.25034 (4) | 0.0101 (3) | |
| H2'A | 0.0559 | 0.2524 | 0.2370 | 0.012* | |
| C3' | 0.2123 (2) | 0.36920 (15) | 0.28278 (4) | 0.0099 (3) | |
| H3' | 0.3721 | 0.3514 | 0.2716 | 0.012* | |
| C4' | 0.1802 (2) | 0.51388 (16) | 0.28752 (4) | 0.0117 (3) | |
| H4' | 0.0287 | 0.5311 | 0.3019 | 0.014* | |
| C5' | 0.3782 (3) | 0.57353 (16) | 0.31172 (4) | 0.0146 (3) | |
| H5'1 | 0.5311 | 0.5562 | 0.2986 | 0.018* | |
| H5'2 | 0.3571 | 0.6687 | 0.3145 | 0.018* | |
| C2 | 0.1186 (2) | 0.18576 (15) | 0.11985 (4) | 0.0131 (3) | |
| H2 | 0.0070 | 0.1209 | 0.1129 | 0.016* | |
| C4 | 0.2533 (2) | 0.34050 (15) | 0.15984 (4) | 0.0105 (3) | |
| C5 | 0.4532 (2) | 0.36319 (16) | 0.13671 (4) | 0.0112 (3) | |
| C6 | 0.4876 (2) | 0.28892 (15) | 0.10027 (4) | 0.0109 (3) | |
| C8 | 0.4676 (2) | 0.49782 (16) | 0.18581 (4) | 0.0125 (3) | |
| H8 | 0.5167 | 0.5658 | 0.2035 | 0.015* | |
| O11 | 0.74791 (17) | 0.46360 (11) | −0.03135 (3) | 0.0157 (2) | |
| O12 | 0.87436 (18) | 0.65877 (12) | −0.01122 (3) | 0.0183 (2) | |
| N11 | 1.3226 (2) | 0.60432 (13) | −0.03067 (3) | 0.0122 (2) | |
| H11A | 1.2980 | 0.6394 | −0.0558 | 0.018* | |
| H11B | 1.4727 | 0.5751 | −0.0289 | 0.018* | |
| H11C | 1.2976 | 0.6660 | −0.0112 | 0.018* | |
| N16 | 1.2090 (2) | 0.64150 (14) | 0.10327 (3) | 0.0135 (2) | |
| H16 | 1.2594 | 0.7041 | 0.1193 | 0.016* | |
| N18 | 0.9928 (2) | 0.48969 (13) | 0.07782 (4) | 0.0128 (2) | |
| H18 | 0.8760 | 0.4350 | 0.0743 | 0.015* | |
| C11 | 0.9065 (2) | 0.54546 (15) | −0.02257 (4) | 0.0114 (3) | |
| C12 | 1.1587 (2) | 0.49425 (15) | −0.02426 (4) | 0.0112 (3) | |
| H12 | 1.1728 | 0.4332 | −0.0479 | 0.013* | |
| C13 | 1.2197 (2) | 0.41984 (15) | 0.01512 (4) | 0.0124 (3) | |
| H13A | 1.1188 | 0.3418 | 0.0171 | 0.015* | |
| H13B | 1.3844 | 0.3901 | 0.0136 | 0.015* | |
| C14 | 1.1884 (2) | 0.50020 (15) | 0.05270 (4) | 0.0114 (3) | |
| C15 | 1.3233 (2) | 0.59562 (16) | 0.06909 (4) | 0.0127 (3) | |
| H15 | 1.4695 | 0.6254 | 0.0589 | 0.015* | |
| C17 | 1.0109 (2) | 0.57579 (16) | 0.10796 (4) | 0.0141 (3) | |
| H17 | 0.9004 | 0.5881 | 0.1293 | 0.017* |
| U11 | U22 | U33 | U12 | U13 | U23 | |
| P1 | 0.00954 (14) | 0.0178 (2) | 0.00845 (13) | 0.00116 (14) | −0.00034 (11) | −0.00065 (12) |
| O3 | 0.0097 (4) | 0.0226 (7) | 0.0127 (4) | 0.0035 (4) | 0.0010 (4) | 0.0021 (4) |
| O5 | 0.0160 (5) | 0.0288 (7) | 0.0100 (4) | −0.0002 (5) | 0.0013 (4) | −0.0004 (4) |
| O2' | 0.0084 (4) | 0.0224 (6) | 0.0118 (4) | 0.0001 (4) | 0.0015 (3) | 0.0029 (4) |
| O3' | 0.0124 (4) | 0.0138 (6) | 0.0086 (4) | −0.0009 (4) | −0.0008 (3) | 0.0008 (3) |
| O4' | 0.0182 (5) | 0.0096 (6) | 0.0111 (4) | 0.0003 (4) | 0.0001 (4) | −0.0003 (3) |
| O5' | 0.0174 (5) | 0.0183 (6) | 0.0124 (4) | 0.0006 (4) | −0.0029 (4) | −0.0045 (4) |
| O6 | 0.0121 (4) | 0.0167 (6) | 0.0134 (4) | 0.0007 (4) | 0.0028 (3) | −0.0009 (4) |
| N1 | 0.0123 (5) | 0.0113 (7) | 0.0123 (5) | 0.0002 (5) | 0.0003 (4) | −0.0020 (4) |
| N3 | 0.0115 (5) | 0.0134 (7) | 0.0137 (5) | −0.0018 (4) | −0.0004 (4) | −0.0002 (4) |
| N7 | 0.0109 (5) | 0.0139 (7) | 0.0132 (5) | −0.0021 (4) | −0.0002 (4) | 0.0001 (4) |
| N9 | 0.0093 (5) | 0.0125 (7) | 0.0096 (4) | −0.0008 (4) | 0.0006 (4) | 0.0006 (4) |
| C1' | 0.0096 (6) | 0.0129 (8) | 0.0103 (5) | 0.0010 (5) | 0.0013 (4) | −0.0011 (4) |
| C2' | 0.0080 (5) | 0.0126 (8) | 0.0096 (5) | −0.0008 (5) | 0.0001 (4) | 0.0004 (5) |
| C3' | 0.0090 (5) | 0.0125 (8) | 0.0083 (5) | 0.0001 (5) | −0.0003 (4) | 0.0006 (4) |
| C4' | 0.0117 (6) | 0.0133 (8) | 0.0101 (5) | 0.0005 (5) | −0.0008 (4) | −0.0012 (5) |
| C5' | 0.0151 (6) | 0.0130 (8) | 0.0157 (6) | −0.0015 (6) | −0.0019 (5) | −0.0024 (5) |
| C2 | 0.0126 (6) | 0.0120 (8) | 0.0146 (5) | −0.0009 (5) | 0.0000 (5) | 0.0009 (5) |
| C4 | 0.0108 (5) | 0.0110 (8) | 0.0097 (5) | 0.0016 (5) | −0.0003 (4) | 0.0018 (4) |
| C5 | 0.0098 (5) | 0.0132 (8) | 0.0107 (5) | 0.0003 (5) | −0.0009 (4) | 0.0022 (5) |
| C6 | 0.0113 (6) | 0.0098 (8) | 0.0116 (5) | 0.0025 (5) | −0.0009 (4) | 0.0021 (5) |
| C8 | 0.0104 (5) | 0.0138 (8) | 0.0132 (6) | −0.0021 (5) | −0.0012 (5) | 0.0005 (5) |
| O11 | 0.0106 (4) | 0.0189 (7) | 0.0175 (5) | 0.0000 (4) | −0.0010 (4) | −0.0040 (4) |
| O12 | 0.0151 (5) | 0.0155 (7) | 0.0244 (5) | 0.0054 (4) | −0.0016 (4) | −0.0069 (4) |
| N11 | 0.0099 (5) | 0.0159 (7) | 0.0107 (5) | 0.0003 (4) | −0.0003 (4) | −0.0013 (4) |
| N16 | 0.0141 (5) | 0.0153 (7) | 0.0112 (5) | −0.0013 (5) | −0.0010 (4) | −0.0005 (4) |
| N18 | 0.0100 (5) | 0.0144 (7) | 0.0141 (5) | −0.0011 (5) | −0.0006 (4) | 0.0016 (4) |
| C11 | 0.0107 (5) | 0.0148 (8) | 0.0087 (5) | 0.0024 (5) | −0.0016 (4) | −0.0010 (4) |
| C12 | 0.0102 (5) | 0.0117 (8) | 0.0118 (5) | 0.0008 (5) | −0.0017 (4) | −0.0023 (5) |
| C13 | 0.0134 (6) | 0.0101 (8) | 0.0138 (5) | 0.0018 (5) | −0.0022 (5) | −0.0005 (5) |
| C14 | 0.0097 (5) | 0.0129 (8) | 0.0116 (5) | 0.0000 (5) | −0.0010 (4) | 0.0026 (5) |
| C15 | 0.0120 (6) | 0.0152 (8) | 0.0111 (5) | −0.0010 (5) | −0.0003 (5) | 0.0021 (5) |
| C17 | 0.0131 (6) | 0.0172 (9) | 0.0122 (5) | 0.0015 (6) | 0.0008 (5) | 0.0019 (5) |
| P1—O3 | 1.4968 (11) | C4'—H4' | 1.00 |
| P1—O5 | 1.4786 (11) | C5'—H5'1 | 0.99 |
| P1—O3' | 1.6258 (11) | C5'—H5'2 | 0.99 |
| P1—O5' | 1.6060 (13) | C2—H2 | 0.95 |
| O2'—C2' | 1.4117 (16) | C4—C5 | 1.3903 (18) |
| O2'—H2' | 0.84 | C5—C6 | 1.4318 (19) |
| O3'—C3' | 1.4309 (15) | C8—H8 | 0.95 |
| O4'—C1' | 1.4232 (17) | O11—C11 | 1.2695 (17) |
| O4'—C4' | 1.4348 (16) | O12—C11 | 1.2370 (19) |
| O5'—C5' | 1.4568 (17) | N11—C12 | 1.4837 (19) |
| O6—C6 | 1.2412 (17) | N11—H11A | 0.91 |
| N1—C2 | 1.3613 (18) | N11—H11B | 0.91 |
| N1—C6 | 1.392 (2) | N11—H11C | 0.91 |
| N1—H1 | 0.88 | N16—C17 | 1.3266 (19) |
| N3—C2 | 1.3100 (18) | N16—C15 | 1.3798 (17) |
| N3—C4 | 1.3555 (19) | N16—H16 | 0.88 |
| N7—C8 | 1.3138 (18) | N18—C17 | 1.3312 (19) |
| N7—C5 | 1.3777 (19) | N18—C14 | 1.3918 (17) |
| N9—C8 | 1.3746 (18) | N18—H18 | 0.88 |
| N9—C4 | 1.3750 (18) | C11—C12 | 1.5339 (19) |
| N9—C1' | 1.4716 (17) | C12—C13 | 1.5409 (19) |
| C1'—C2' | 1.544 (2) | C12—H12 | 1.00 |
| C1'—H1' | 1.00 | C13—C14 | 1.4945 (19) |
| C2'—C3' | 1.5204 (18) | C13—H13A | 0.99 |
| C2'—H2'A | 1.00 | C13—H13B | 0.99 |
| C3'—C4' | 1.507 (2) | C14—C15 | 1.358 (2) |
| C3'—H3' | 1.00 | C15—H15 | 0.95 |
| C4'—C5' | 1.511 (2) | C17—H17 | 0.95 |
| O3—P1—O5 | 117.75 (6) | N1—C2—H2 | 117.3 |
| O3—P1—O3' | 108.67 (6) | N3—C4—N9 | 126.36 (12) |
| O3—P1—O5' | 110.00 (6) | N3—C4—C5 | 128.42 (13) |
| O3'—P1—O5 | 107.58 (6) | N9—C4—C5 | 105.21 (12) |
| O3'—P1—O5' | 103.48 (6) | N7—C5—C4 | 111.00 (12) |
| O5—P1—O5' | 108.40 (6) | N7—C5—C6 | 130.39 (12) |
| C2'—O2'—H2' | 109.5 | C4—C5—C6 | 118.58 (13) |
| C3'—O3'—P1 | 111.42 (8) | O6—C6—N1 | 120.68 (13) |
| C1'—O4'—C4' | 108.09 (11) | O6—C6—C5 | 128.30 (14) |
| C5'—O5'—P1 | 119.39 (9) | N1—C6—C5 | 111.00 (12) |
| C2—N1—C6 | 125.17 (12) | N7—C8—N9 | 113.24 (13) |
| C2—N1—H1 | 117.4 | N7—C8—H8 | 123.4 |
| C6—N1—H1 | 117.4 | N9—C8—H8 | 123.4 |
| C2—N3—C4 | 111.32 (12) | C12—N11—H11A | 109.5 |
| C8—N7—C5 | 104.27 (12) | C12—N11—H11B | 109.5 |
| C8—N9—C4 | 106.28 (11) | H11A—N11—H11B | 109.5 |
| C8—N9—C1' | 126.53 (12) | C12—N11—H11C | 109.5 |
| C4—N9—C1' | 126.45 (11) | H11A—N11—H11C | 109.5 |
| O4'—C1'—N9 | 106.50 (11) | H11B—N11—H11C | 109.5 |
| O4'—C1'—C2' | 107.57 (10) | C17—N16—C15 | 108.84 (13) |
| N9—C1'—C2' | 113.35 (12) | C17—N16—H16 | 125.6 |
| O4'—C1'—H1' | 109.8 | C15—N16—H16 | 125.6 |
| N9—C1'—H1' | 109.8 | C17—N18—C14 | 109.00 (12) |
| C2'—C1'—H1' | 109.8 | C17—N18—H18 | 125.5 |
| O2'—C2'—C3' | 113.65 (10) | C14—N18—H18 | 125.5 |
| O2'—C2'—C1' | 105.27 (11) | O12—C11—O11 | 125.94 (13) |
| C3'—C2'—C1' | 97.92 (11) | O12—C11—C12 | 118.28 (12) |
| O2'—C2'—H2'A | 112.9 | O11—C11—C12 | 115.70 (13) |
| C3'—C2'—H2'A | 112.9 | N11—C12—C11 | 109.58 (12) |
| C1'—C2'—H2'A | 112.9 | N11—C12—C13 | 110.80 (11) |
| O3'—C3'—C4' | 110.82 (10) | C11—C12—C13 | 110.62 (11) |
| O3'—C3'—C2' | 116.82 (11) | N11—C12—H12 | 108.6 |
| C4'—C3'—C2' | 101.13 (11) | C11—C12—H12 | 108.6 |
| O3'—C3'—H3' | 109.2 | C13—C12—H12 | 108.6 |
| C4'—C3'—H3' | 109.2 | C14—C13—C12 | 112.93 (12) |
| C2'—C3'—H3' | 109.2 | C14—C13—H13A | 109.0 |
| O4'—C4'—C3' | 103.32 (10) | C12—C13—H13A | 109.0 |
| O4'—C4'—C5' | 114.48 (12) | C14—C13—H13B | 109.0 |
| C3'—C4'—C5' | 111.33 (12) | C12—C13—H13B | 109.0 |
| O4'—C4'—H4' | 109.2 | H13A—C13—H13B | 107.8 |
| C3'—C4'—H4' | 109.2 | C15—C14—N18 | 106.06 (12) |
| C5'—C4'—H4' | 109.2 | C15—C14—C13 | 131.17 (13) |
| O5'—C5'—C4' | 104.02 (12) | N18—C14—C13 | 122.74 (13) |
| O5'—C5'—H5'1 | 111.0 | C14—C15—N16 | 107.50 (12) |
| C4'—C5'—H5'1 | 111.0 | C14—C15—H15 | 126.2 |
| O5'—C5'—H5'2 | 111.0 | N16—C15—H15 | 126.2 |
| C4'—C5'—H5'2 | 111.0 | N16—C17—N18 | 108.59 (12) |
| H5'1—C5'—H5'2 | 109.0 | N16—C17—H17 | 125.7 |
| N3—C2—N1 | 125.45 (14) | N18—C17—H17 | 125.7 |
| N3—C2—H2 | 117.3 | ||
| O3—P1—O3'—C3' | −67.24 (10) | C8—N9—C4—N3 | −179.69 (14) |
| O5—P1—O3'—C3' | 164.26 (9) | C1'—N9—C4—N3 | −9.1 (2) |
| O5'—P1—O3'—C3' | 49.65 (10) | C8—N9—C4—C5 | 0.73 (15) |
| O3—P1—O5'—C5' | 63.87 (11) | C1'—N9—C4—C5 | 171.35 (13) |
| O5—P1—O5'—C5' | −166.11 (10) | C8—N7—C5—C4 | −0.18 (16) |
| O3'—P1—O5'—C5' | −52.08 (11) | C8—N7—C5—C6 | 177.46 (15) |
| C4'—O4'—C1'—N9 | −118.53 (11) | N3—C4—C5—N7 | −179.93 (14) |
| C4'—O4'—C1'—C2' | 3.29 (13) | N9—C4—C5—N7 | −0.36 (16) |
| C8—N9—C1'—O4' | −5.89 (18) | N3—C4—C5—C6 | 2.1 (2) |
| C4—N9—C1'—O4' | −174.66 (12) | N9—C4—C5—C6 | −178.31 (12) |
| C8—N9—C1'—C2' | −123.97 (14) | C2—N1—C6—O6 | 179.53 (13) |
| C4—N9—C1'—C2' | 67.26 (17) | C2—N1—C6—C5 | −2.06 (19) |
| O4'—C1'—C2'—O2' | 87.45 (12) | N7—C5—C6—O6 | 0.8 (3) |
| N9—C1'—C2'—O2' | −155.09 (10) | C4—C5—C6—O6 | 178.25 (14) |
| O4'—C1'—C2'—C3' | −29.79 (12) | N7—C5—C6—N1 | −177.49 (14) |
| N9—C1'—C2'—C3' | 87.66 (12) | C4—C5—C6—N1 | 0.00 (18) |
| P1—O3'—C3'—C4' | −60.89 (12) | C5—N7—C8—N9 | 0.68 (16) |
| P1—O3'—C3'—C2' | −175.96 (10) | C4—N9—C8—N7 | −0.92 (16) |
| O2'—C2'—C3'—O3' | 53.48 (17) | C1'—N9—C8—N7 | −171.53 (13) |
| C1'—C2'—C3'—O3' | 164.04 (11) | O12—C11—C12—N11 | −26.71 (16) |
| O2'—C2'—C3'—C4' | −66.87 (14) | O11—C11—C12—N11 | 156.47 (11) |
| C1'—C2'—C3'—C4' | 43.69 (11) | O12—C11—C12—C13 | 95.75 (15) |
| C1'—O4'—C4'—C3' | 25.38 (13) | O11—C11—C12—C13 | −81.08 (15) |
| C1'—O4'—C4'—C5' | 146.61 (12) | N11—C12—C13—C14 | 64.69 (15) |
| O3'—C3'—C4'—O4' | −168.80 (10) | C11—C12—C13—C14 | −57.05 (15) |
| C2'—C3'—C4'—O4' | −44.26 (12) | C17—N18—C14—C15 | 0.05 (16) |
| O3'—C3'—C4'—C5' | 67.87 (14) | C17—N18—C14—C13 | −178.10 (13) |
| C2'—C3'—C4'—C5' | −167.60 (11) | C12—C13—C14—C15 | −77.04 (19) |
| P1—O5'—C5'—C4' | 58.05 (14) | C12—C13—C14—N18 | 100.61 (16) |
| O4'—C4'—C5'—O5' | −178.00 (11) | N18—C14—C15—N16 | −0.29 (16) |
| C3'—C4'—C5'—O5' | −61.29 (14) | C13—C14—C15—N16 | 177.65 (14) |
| C4—N3—C2—N1 | −0.3 (2) | C17—N16—C15—C14 | 0.43 (16) |
| C6—N1—C2—N3 | 2.4 (2) | C15—N16—C17—N18 | −0.40 (16) |
| C2—N3—C4—N9 | 178.59 (14) | C14—N18—C17—N16 | 0.22 (16) |
| C2—N3—C4—C5 | −1.9 (2) |
| D—H···A | D—H | H···A | D···A | D—H···A |
| O2′—H2′···O3′ | 0.84 | 2.56 | 2.9671 (15) | 111 |
| O2′—H2′···O3i | 0.84 | 1.96 | 2.7340 (14) | 154 |
| N1—H1···O11ii | 0.88 | 1.83 | 2.6931 (17) | 166 |
| N11—H11A···O5iii | 0.91 | 1.79 | 2.6906 (16) | 168 |
| N11—H11B···O11iv | 0.91 | 1.95 | 2.8266 (17) | 162 |
| N11—H11B···O12iv | 0.91 | 2.52 | 3.2622 (19) | 140 |
| N11—H11C···O12v | 0.91 | 2.00 | 2.8125 (18) | 148 |
| N16—H16···O3vi | 0.88 | 1.87 | 2.7370 (17) | 169 |
| N18—H18···O6 | 0.88 | 1.92 | 2.7886 (18) | 169 |
| C2′—H2′A···O4′vii | 1.00 | 2.41 | 3.082 (2) | 124 |
| C3′—H3′···O2′iv | 1.00 | 2.44 | 3.400 (2) | 160 |
| C12—H12···O6viii | 1.00 | 2.53 | 3.428 (2) | 149 |
| C17—H17···N7 | 0.95 | 2.35 | 3.079 (2) | 133 |
| Symmetry codes: (i) x−1, y, z; (ii) x−1/2, −y+1/2, −z; (iii) −x+3/2, −y+1, z−1/2; (iv) x+1, y, z; (v) x+1/2, −y+3/2, −z; (vi) −x+2, y+1/2, −z+1/2; (vii) −x, y−1/2, −z+1/2; (viii) x+1/2, −y+1/2, −z. |
Experimental details
| Crystal data | |
| Chemical formula | C6H10N3O2+·C10H10N4O7P− |
| Mr | 485.36 |
| Crystal system, space group | Orthorhombic, P212121 |
| Temperature (K) | 90 |
| a, b, c (Å) | 5.709 (2), 10.284 (3), 32.795 (9) |
| V (Å3) | 1925.4 (10) |
| Z | 4 |
| Radiation type | Mo Kα |
| µ (mm−1) | 0.22 |
| Crystal size (mm) | 0.12 × 0.07 × 0.04 |
| Data collection | |
| Diffractometer | Oxford Xcalibur PX κ-geometry diffractometer with an Onyx CCD camera |
| Absorption correction | Multi-scan (CrysAlis RED; Oxford Diffraction, 2009) |
| Tmin, Tmax | 0.949, 1.000 |
| No. of measured, independent and observed [I > 2σ(I)] reflections | 17935, 5114, 4778 |
| Rint | 0.032 |
| (sin θ/λ)max (Å−1) | 0.703 |
| Refinement | |
| R[F2 > 2σ(F2)], wR(F2), S | 0.030, 0.077, 1.04 |
| No. of reflections | 5114 |
| No. of parameters | 300 |
| H-atom treatment | H-atom parameters constrained |
| Δρmax, Δρmin (e Å−3) | 0.35, −0.27 |
| Absolute structure | From known absolute configuration and anomalous dispersion effects (Flack, 1983), with 1877 Friedel pairs |
| Absolute structure parameter | 0.20 (6) |
Computer programs: CrysAlis CCD (Oxford Diffraction, 2009), CrysAlis RED (Oxford Diffraction, 2009), SHELXS97 (Sheldrick, 2008), SHELXL97 (Sheldrick, 2008), DIAMOND (Brandenburg, 2005).
| P1—O3 | 1.4968 (11) | P1—O5' | 1.6060 (13) |
| P1—O5 | 1.4786 (11) | O11—C11 | 1.2695 (17) |
| P1—O3' | 1.6258 (11) | O12—C11 | 1.2370 (19) |
| O3—P1—O5 | 117.75 (6) | O3'—P1—O5 | 107.58 (6) |
| O3—P1—O3' | 108.67 (6) | O3'—P1—O5' | 103.48 (6) |
| O3—P1—O5' | 110.00 (6) | O5—P1—O5' | 108.40 (6) |
| O3—P1—O3'—C3' | −67.24 (10) | P1—O3'—C3'—C2' | −175.96 (10) |
| O5—P1—O3'—C3' | 164.26 (9) | P1—O5'—C5'—C4' | 58.05 (14) |
| O5'—P1—O3'—C3' | 49.65 (10) | O4'—C4'—C5'—O5' | −178.00 (11) |
| O3—P1—O5'—C5' | 63.87 (11) | C3'—C4'—C5'—O5' | −61.29 (14) |
| O5—P1—O5'—C5' | −166.11 (10) | O11—C11—C12—N11 | 156.47 (11) |
| O3'—P1—O5'—C5' | −52.08 (11) | N11—C12—C13—C14 | 64.69 (15) |
| C4—N9—C1'—O4' | −174.66 (12) | C11—C12—C13—C14 | −57.05 (15) |
| P1—O3'—C3'—C4' | −60.89 (12) | C12—C13—C14—N18 | 100.61 (16) |
| D—H···A | D—H | H···A | D···A | D—H···A |
| O2'—H2'···O3' | 0.84 | 2.56 | 2.9671 (15) | 111 |
| O2'—H2'···O3i | 0.84 | 1.96 | 2.7340 (14) | 154 |
| N1—H1···O11ii | 0.88 | 1.83 | 2.6931 (17) | 166 |
| N11—H11A···O5iii | 0.91 | 1.79 | 2.6906 (16) | 168 |
| N11—H11B···O11iv | 0.91 | 1.95 | 2.8266 (17) | 162 |
| N11—H11B···O12iv | 0.91 | 2.52 | 3.2622 (19) | 140 |
| N11—H11C···O12v | 0.91 | 2.00 | 2.8125 (18) | 148 |
| N16—H16···O3vi | 0.88 | 1.87 | 2.7370 (17) | 169 |
| N18—H18···O6 | 0.88 | 1.92 | 2.7886 (18) | 169 |
| C2'—H2'A···O4'vii | 1.00 | 2.41 | 3.082 (2) | 124 |
| C3'—H3'···O2'iv | 1.00 | 2.44 | 3.400 (2) | 160 |
| C12—H12···O6viii | 1.00 | 2.53 | 3.428 (2) | 149 |
| C17—H17···N7 | 0.95 | 2.35 | 3.079 (2) | 133 |
| Symmetry codes: (i) x−1, y, z; (ii) x−1/2, −y+1/2, −z; (iii) −x+3/2, −y+1, z−1/2; (iv) x+1, y, z; (v) x+1/2, −y+3/2, −z; (vi) −x+2, y+1/2, −z+1/2; (vii) −x, y−1/2, −z+1/2; (viii) x+1/2, −y+1/2, −z. |




High-resolution crystal structures of amino acid–nucleotide complexes can serve as model systems to investigate protein–nucleic acid interactions and their manner of recognition. Nucleic acid-binding proteins discriminate between potential binding sites based on their sequence or structure or a combination of both. The forces involved in noncovalent protein–nucleotide/nucleic acid complex formation are relatively weak (hydrogen bonds, hydrophobic interactions, and π–π stacking between the base and aromatic side chain), but still crucial for the stability of the complex, and the overall affinity results from the sum of many interactions. Electrostatics, hydrophobic interactions and polar contacts involving direct or water-mediated hydrogen bonds are all evident and widespread (Rice & Correll, 2008).
In DNA–protein complexes, histidine has been shown to prefer interactions with guanine (Luscombe et al., 2001), which is a 2-amino derivative of hypoxanthine and thus has the same Hoogsteen edge crucial for protein–DNA recognition. The nucleotide Hoogsteen edge, with its unique functional groups (hydrogen-bond donors and acceptors) exposed in the major groove, is associated with the main specifity of protein–B-DNA complexes. This is distinct from protein–RNA and protein–nucleotide recognition, in which the different edges of a base may play a similar role in protein binding: the Watson–Crick edge, the Hoogsteen edge and the sugar edge (Leontis & Westhof, 2001).
The only examples of coordinating-metal-free complexes of amino acid/oligopeptide–nucleotide deposited so far in the Cambridge Structural Database (CSD, Version?; Allen, 2002) are three isomorphous complexes of IMP with L-Ser (CSD refcode ZUWQEN; Mukhopadhyay et al., 1995), L-Glu (QUSMIA; Bhattacharya et al., 2000) and L-Gln (LIRQUY; Bera et al., 1999), two complexes of N7-methylguanosine 5'-monophosphate (m7GMP) with L-Phe and Trp–Glu [DUMJEA10 (Ishida et al., 1988) and SEKXIP10 (Ishida et al., 1991), respectively], and the most recent isomorphous L-His–IMP.MeOH.H2O (TUVZIU; Ślepokura & Petrus, 2010) and L-His–IMP.2.5H2O (Ślepokura & Makarewicz, 2012). No crystal data on cyclic nucleotide–amino acid complexes are available to date. The analysis of structural details is difficult in the first four of the above-mentioned structures, mainly due to the lack of at least some of the H atoms and therefore incorrect ionization states. In addition, the crystal structure of the highly hydrated IMP–Ser/Glu/Gln complexes is dominated by IMP–IMP interactions and no direct specific nucleobase–amino acid functional side [secondary?] interactions are observed.
Therefore, the synthesis and structural analysis of the title L-His–cIMP complex, (I), which reveals both specific and nonspecific interactions of L-histidine (His) with inosine 3':5'-cyclic monophosphate (cIMP), have been undertaken. The conformation and binding mode of cIMP are compared with those found in the solvated L-His–IMP complexes and the other amino acid–IMP complexes deposited in the CSD, as well as with cIMP crystal structures reported to date, viz. cIMP.H2O (BEPRAP; Sundaralingam et al., 1982) and [Cu(cIMP)(phenanthroline)(H2O)2]NO3.2H2O (CATTEW; Sheldrick, 1983).
The asymmetric unit of (I) consists of a cIMP monoanion and a His cation, as shown in Fig. 1. Analogous to IMP in two isomorphous solvated L-His–IMP complexes, the cIMP nucleotide in (I) adopts a typical anti conformation about the N-glycosidic C1'—N9 bond, although with a χCN O4'—C1'—N9—C4 torsion angle of -174.66 (12)°, which is uncommon [compared with -139.2 (5) and -138.5 (3)° observed in IMP in its complexes]. As shown in Fig. 2, the orientation of the Hyp base relative to the ribose ring in IMP and cIMP in their complexes with L-His is the main difference in the structures of these analogous nucleotides, despite the same formal anti glycosyl conformation.
Analogous to IMP in the solvated L-His–IMP complexes, the cIMP nucleotide in (I) is puckered in an envelope C3'-endo (3E) manner, which is confirmed by the pseudorotation parameters (Rao et al., 1981) P = 14.7 (1)° and τm = 46.5 (1)°, and the Cremer & Pople (1975) puckering parameters q2 = 0.4561 (15) Å and ϕ2 = 283.22 (17)°. It is worth mentioning that the 3E conformation is one of two typically observed in free IMP and other free and protein-bound nucleotides (Allen, 2002; Moodie & Thornton, 1993), and was also observed in the solvated L-His–IMP complexes, but not in the other previously reported amino acid–nucleotide (both IMP and m7GMP) complexes, in which C2'-endo (2E) puckering is the most common. However, the ribose in the only crystal structures of cIMP reported so far, viz. cIMP.H2O (BEPRAP; Sundaralingam et al., 1982) and [Cu(cIMP)(phenanthroline)(H2O)2]NO3.2H2O (CATTEW; Sheldrick, 1983), is found in a different puckering conformation, i.e. C3'-endo/C4'-exo (3T4). The ribose in protein-bound IMP does not reveal any conformational preference either.
The chair (C) conformation of the six-membered O/P/O/C/C/C 1,3,2-dioxaphosphorinane ring of the cIMP anion is significantly flattened at the P atom. This deformation of the ring towards an envelope (E) is reflected in the values of the dihedral angles between the least-squares plane through the four central atoms of the ring (O3'/O5'/C3'/C5') and the O3'/P1/O5' and C3'/C4'/C5' planes [ϕ1 = 44.5 (1)° and ϕ2 = 59.4 (1)°; ϕ2 - ϕ1 = 14.9 (2)°], as well as in the Cremer–Pople puckering parameters: Q = 0.5959 (13) Å, θ = 172.88 (12)° and ϕ = 346.0 (11)°. It has been shown that, in small six-membered cyclic phosphates, the extent of C→ E deformation is related to protonation or esterification of the cyclic phosphate group (Ślepokura & Lis, 2004; Ślepokura, 2008; Ślepokura & Mitaszewska, 2011). It is convenient to use the values of |ϕ2 - ϕ1| (determining the dihedral angles between the O—P—O and C—C—C planes) for a quantitative description of ring flattening. In small cyclic phosphates, the |ϕ2 - ϕ1| values amount to several degrees for compounds with an ionized phosphate group, and to more than 12° for acidic or esterified molecules. However, this general rule seems not to be true in the cIMP anion of (I) presented here, in which ϕ2 - ϕ1 = 14.9 (2)°, nor in other cIMP anions reported to date (ϕ2 - ϕ1 = 18.2 and 14.8° for BEPRAP and CATTEW, respectively), nor in the other 3':5'-cyclic nucleotides deposited in the CSD, in which ϕ2 - ϕ1 ranges from 10.9 to 20.9° (Allen, 2008).
Selected geometric parameters for (I), given in Table 1, indicate the deformation of the phosphate group from the ideal tetrahedral shape, which is seen particularly in the endocyclic O3'—P1—O5' and exocyclic O3—P1—O5 bond angles, which are, respectively, the smallest [103.48 (6)°] and the largest [117.75 (6)°). These values of the endo- and exocyclic O—P—O angles correlate with the respective P—O bond lengths. In (I), as in other ionized six-membered cyclic phosphates (Ślepokura, 2008; Ślepokura & Mitaszewska, 2011), the P—Oendo bonds are all approximately 1.6 Å, and roughly 0.1 Å longer than the P—Oexo bonds.
The L-His cation in its complexes with both IMP and cIMP has the carboxyl group deprotonated, and amino atom N11 and imidazole atom N16 (Nε) protonated. As seen in Fig. 2(a), the His conformation is different. In both cases, the planes defined by the imidazolium ring and by the backbone NH3+—Cα12—COO- fragment are both approximately perpendicular to the Cα12—Cβ13—Cγ14 plane, which is reflected in the interplanar angles of 78.4 (1) and 87.6 (1)°, respectively, in (I), compared with 86.8 (4)/87.7 (4)° and 88.4 (2)/88.3 (2)° in two solvated L-His–IMP complexes (Ślepokura & Petrus, 2010; Ślepokura & Makarewicz, 2012). The location of the protonated atoms N11 and N18 (Nδ) on different sides of the His cation in (I) differentiates it from His in complexes with IMP, and plays a role in the His···cIMP and His···IMP interactions, as will be discussed below. As shown in Fig. 2, and confirmed by the pseudotorsion angles N11—Cα12—Cγ14—Cδ15 of 2.7 (1)° [for His in (I)] and N11—Cα12—Cγ14—Nδ18 of -11.2 (1) and -10.6 (3)° (for His in IMP complexes), in L-His–cIMP the carboxylate (COO-) group and the protonated N18 atom (Nδ) are located on the same side of the His cation, whereas in the solvated L-His–IMP complexes, the protonated atoms N11 (NH3+) and N18 (Nδ) are on the same side of the His cation.
All of the differences noted above in the chemical nature and structure of the components of His–IMP and His–cIMP complexes translate into the nature of the interactions between His cations (His···His), between nucleotide anions (cIMP···cIMP) and, the most interesting and crucial to the amino acid–nucleotide recognition process, the His···cIMP contacts. As shown in Fig. 3, both His···His and cIMP···cIMP interactions arrange adjacent His cations or cIMP anions into infinite ribbons running down the a axis. The overall architecture of the His ribbon in (I) is similar to that in the solvated L-His–IMP complexes: each His cation is joined to four adjacent [What? Text missing] (related by a direct a-axis translation and a 21 symmetry operation) via a network of bifurcated N–H···O hydrogen bonds of a salt-bridge type, which are however much more compact than in His–IMP complexes (Fig. 3a and Table 2). As in His–IMP complexes, the imidazolium rings, which are located outside the ribbon, are exposed to specific amino acid–nucleotide interactions (Fig. 3b).
In contrast, the interactions between the cIMP anions in (I) are much less numerous than the IMP···IMP contacts in solvated His–IMP complexes. In (I), a direct a-axis translation generates infinite ribbons of cIMP anions via O—H···O interactions linking the ribosyl O2'—H2' group with atom O3 of an adjacent cyclic phosphate group, and via C—H···O contacts linking the same hydroxyl group with an adjacent ribosyl C atom (Fig. 3b). Notably, the hypoxanthine bases in (I) are not involved in the cIMP···cIMP contacts, and therefore may be used more efficiently for His–cIMP recognition. In addition, the monoionized cyclic phosphate group in (I) has no H atom, which excludes the possibility of phosphate—phosphate interactions. All these observations result in significant differences in mutual orientation and internucleotide interactions between IMP and its cyclic analogue, cIMP in (I). It is worth mentioning that internucleotide contacts seem to predominate in the crystal structures of previously reported nucleotide–amino acid complexes. They are realised mainly via sugar–phosphate interactions (all but His–IMP) and contacts involving the base: sugar–base, like O2'···N3(Hyp) in His–IMP complexes, and base–phosphate, like N1(Hyp)···OP in three other IMP complexes. In Phe–m7GMP and solvated His–IMP complexes, additional phosphate–phosphate interactions also exist. Against this background, the internucleotide interactions in (I), with the only sugar–phosphate contacts present, appear to be very simple.
In both the His–cIMP, (I), and His–IMP crystal complexes, the nucleotide Hoogsteen edge, commonly used in protein–nucleic acid recognition, is also involved in amino acid–nucleotide (His···cIMP and His···IMP) interactions. As shown in Figs. 1 and 3(b), in (I) the histidine side chain–Hyp base recognition is realised via specific edge-to-edge hydrogen bonds utilizing both Hyp Hoogsteen atoms N7 and O6, i.e. Nδ18–H18···O6(Hyp) and Cε17–H17···N7(Hyp) (Table 2), as well as via π–π stacking of the cation···π type [centroid-to-centroid distances of 3.409 (2) and 3.438 (2) Å, respectively, for Cg(His)···Cg(Hyp-six-membered) and Cg(His)···Cg(Hyp-five-membered), with an interplanar spacing of 3.22 (1) Å; Fig. 3b]. In the isomorphous His–IMP complexes, Cδ15–H15···O6(Hyp) and Nε16–H16···N7(Hyp) interactions are found. Different His–Hoogsteen(Hyp) hydrogen bonds in His–cIMP and His–IMP complexes are accompanied by different conformations of the His cation (discussed above) and thus different orientations of the imidazolium ring relative to the nucleotide base. However, in each case, the orientation is retained such that the interactions of the δ and ε positions of the His cation with, respectively, atoms O6(Hyp) and N7(Hyp) of the nucleotide anion are achieved.
The specific nucleotide–amino acid interactions involving the base Hoogsteen edge are crucial in the recognition process. Direct amino acid side chain–Hyp interactions were not observed in the other IMP–amino acid complexes, except for His-IMP, but they may be analysed in macromolecular protein–IMP complexes (examples in Ślepokura & Petrus, 2010). However, it is also known that, in protein–IMP complexes, the Watson–Crick edge is also involved in nucleotide–amino acid interactions, e.g. N1(Hyp)···amino acid side-chain interactions are commonly observed. As shown in Fig. 4, in (I) each cIMP anion utilizes the base Watson–Crick edge to form nonspecific N—H···O and C—H···O His···cIMP interactions, linking the Hyp atom N1 to the carboxyl group of the adjacent His cation, and Hyp atom O6 to the C12—H12 group of the same His cation. In this way, two different R22(8) rings (Bernstein et al., 1995) are generated for each Hyp base with the participation of two different His cations. In contrast, the Watson–Crick edge of IMP in His-IMP complexes is involved in solvent-mediated IMP···MeOH/H2O···IMP contacts, and therefore does not bind His at all.
As seen from a number of contacts involving the cIMP nucleotide anionic cyclic phosphate group in (I), it may be stated that its role in nucleotide–amino acid interactions is much smaller. The phosphate group, as it has no H atom, forms only two hydrogen bonds: one of a salt-bridge type with ammonium atom N11, and the other with imidazolium atom Nε16 of a different His cation (Fig. 4 and Table 2). As a result of these interactions, each cIMP anion is bound to four different His cations: to two via the nucleotide base and to the other two via the cyclic phosphate group. The fifth His cation is joined by π–π stacking of the cation···π type, as shown in Figs. 3(b) and 4. A similar observation is noted in His–IMP complexes. However, despite the same number of surrounding ions, the nature of these interactions is significantly different. In His–IMP complexes, as many as three of the nearest His cations are bound to the IMP anion via the phosphate group (two of them in a monodentate manner and the third in a bidentate manner).
In conclusion, this paper reports the preparation and solid-state structure of and intermolecular interactions in the first example of a 3':5'-cyclic nucleotide–amino acid complex, namely L-His–cIMP, (I). By comparison with the analogous L-His–IMP complex, it has been demonstrated that the 3':5'-cyclization of the nucleotide may reduce the nucleotide–nucleotide interactions and therefore intensify the nucleotide–amino acid interactions. In (I), both the Hoogsteen edge and the face of the Hyp base participate in specific His···cIMP recognition via edge-to-edge interactions and π–π stacking. The cIMP base Watson–Crick edge is involved in additional nonspecific nucleotide–amino acid contacts, which were not observed in L-His–IMP complexes. In contrast, the role of the phosphate group in nonspecific interactions is reduced compared with His–IMP complexes. This report has also focused on the cIMP conformation, against the background of the available structural information on 3':5'-cyclic nucleotides.