

 journal menu
journal menu organic compounds
organic compounds







 O hydrogen bonds. The overall aggregation pattern is distinctly different from that observed in the glycine–oxalic acid complex.
O hydrogen bonds. The overall aggregation pattern is distinctly different from that observed in the glycine–oxalic acid complex.Supporting information
 | Crystallographic Information File (CIF) https://doi.org/10.1107/S1600536801010042/ob6056sup1.cif |
 | Structure factor file (CIF format) https://doi.org/10.1107/S1600536801010042/ob6056Isup2.hkl |
CCDC reference: 170778
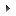

Crystals of (I) were grown from a saturated aqueous solution containing L-alanine and oxalic acid in a stoichiometric ratio.
The absolute structure of (I) was not established by the analysis but is known from the configuration of the starting reagents. The H atoms were placed at calculated positions and were allowed to ride on their respective parent atoms with HFIX instructions using SHELXL97 (Sheldrick, 1997) defaults.
Data collection: CAD-4 Software (Enraf-Nonius, 1989); cell refinement: CAD-4 Software; data reduction: CAD-4 Software; program(s) used to solve structure: SHELXS97 (Sheldrick, 1990); program(s) used to refine structure: SHELXL97 (Sheldrick, 1997); molecular graphics: PLATON (Spek, 1999); software used to prepare material for publication: SHELXL97.
![[Figure 1]](http://journals.iucr.org/e/issues/2001/07/00/ob6056/ob6056fig1thm.gif)
| Fig. 1. The molecular structure of with the atom-numbering scheme and 50% probability displacement ellipsoids. |
| C3H8NO2+·C2HO4− | Dx = 1.490 Mg m−3 Dm = 1.49 Mg m−3 Dm measured by flotation in a mixture of carbon tetrachloride and bromoform |
| Mr = 179.13 | Cu Kα radiation, λ = 1.54180 Å |
| Orthorhombic, P212121 | Cell parameters from 25 reflections |
| a = 5.6304 (15) Å | θ = 4–68° |
| b = 7.2353 (15) Å | µ = 1.23 mm−1 |
| c = 19.597 (3) Å | T = 293 K |
| V = 798.3 (2) Å3 | Needle, colourless |
| Z = 4 | 0.2 × 0.15 × 0.11 mm |
| F(000) = 376 |
| Enraf-Nonius sealed tube diffractometer | 853 reflections with I > 2σ(I) |
| Radiation source: fine-focus sealed tube | Rint = 0.000 |
| Graphite monochromator | θmax = 67.9°, θmin = 4.5° |
| ω–2θ scans | h = 0→6 |
| Absorption correction: ψ scan (North et al., 1968) | k = 0→8 |
| Tmin = 0.79, Tmax = 0.87 | l = 0→23 |
| 879 measured reflections | 2 standard reflections every 25 reflections |
| 879 independent reflections | intensity decay: 0.1% |
| Refinement on F2 | Hydrogen site location: inferred from neighbouring sites |
| Least-squares matrix: full | H-atom parameters constrained |
| R[F2 > 2σ(F2)] = 0.030 | w = 1/[σ2(Fo2) + (0.0481P)2 + 0.1739P] where P = (Fo2 + 2Fc2)/3 |
| wR(F2) = 0.083 | (Δ/σ)max < 0.001 |
| S = 1.09 | Δρmax = 0.14 e Å−3 |
| 879 reflections | Δρmin = −0.14 e Å−3 |
| 112 parameters | Extinction correction: SHELXL97, Fc*=kFc[1+0.001xFc2λ3/sin(2θ)]-1/4 |
| 0 restraints | Extinction coefficient: 0.040 (3) |
| Primary atom site location: structure-invariant direct methods | Absolute structure: see text |
| Secondary atom site location: difference Fourier map |
| C3H8NO2+·C2HO4− | V = 798.3 (2) Å3 |
| Mr = 179.13 | Z = 4 |
| Orthorhombic, P212121 | Cu Kα radiation |
| a = 5.6304 (15) Å | µ = 1.23 mm−1 |
| b = 7.2353 (15) Å | T = 293 K |
| c = 19.597 (3) Å | 0.2 × 0.15 × 0.11 mm |
| Enraf-Nonius sealed tube diffractometer | 853 reflections with I > 2σ(I) |
| Absorption correction: ψ scan (North et al., 1968) | Rint = 0.000 |
| Tmin = 0.79, Tmax = 0.87 | 2 standard reflections every 25 reflections |
| 879 measured reflections | intensity decay: 0.1% |
| 879 independent reflections |
| R[F2 > 2σ(F2)] = 0.030 | 0 restraints |
| wR(F2) = 0.083 | H-atom parameters constrained |
| S = 1.09 | Δρmax = 0.14 e Å−3 |
| 879 reflections | Δρmin = −0.14 e Å−3 |
| 112 parameters | Absolute structure: see text |
Geometry. All e.s.d.'s (except the e.s.d. in the dihedral angle between two l.s. planes) are estimated using the full covariance matrix. The cell e.s.d.'s are taken into account individually in the estimation of e.s.d.'s in distances, angles and torsion angles; correlations between e.s.d.'s in cell parameters are only used when they are defined by crystal symmetry. An approximate (isotropic) treatment of cell e.s.d.'s is used for estimating e.s.d.'s involving l.s. planes. |
Refinement. Refinement of F2 against ALL reflections. The weighted R-factor wR and goodness of fit S are based on F2, conventional R-factors R are based on F, with F set to zero for negative F2. The threshold expression of F2 > σ(F2) is used only for calculating R-factors(gt) etc. and is not relevant to the choice of reflections for refinement. R-factors based on F2 are statistically about twice as large as those based on F, and R- factors based on ALL data will be even larger. |
| x | y | z | Uiso*/Ueq | ||
| N1 | 0.2644 (3) | 0.8562 (2) | 0.10946 (7) | 0.0359 (4) | |
| H1A | 0.1867 | 0.8789 | 0.1481 | 0.054* | |
| H1B | 0.4196 | 0.8708 | 0.1163 | 0.054* | |
| H1C | 0.2356 | 0.7408 | 0.0962 | 0.054* | |
| O1 | 0.1991 (3) | 0.9924 (3) | −0.06326 (7) | 0.0511 (5) | |
| H1 | 0.2798 | 0.9719 | −0.0973 | 0.077* | |
| O2 | 0.5153 (3) | 0.8816 (2) | −0.00816 (7) | 0.0411 (4) | |
| O3 | 0.4157 (3) | 0.5324 (4) | 0.21396 (8) | 0.0623 (7) | |
| H3 | 0.5443 | 0.5063 | 0.2311 | 0.093* | |
| O4 | 0.2719 (3) | 0.4667 (3) | 0.31722 (7) | 0.0489 (5) | |
| O5 | −0.0096 (3) | 0.5505 (2) | 0.16329 (6) | 0.0410 (4) | |
| O6 | −0.1695 (3) | 0.4814 (4) | 0.26207 (8) | 0.0760 (8) | |
| C1 | 0.3174 (3) | 0.9450 (3) | −0.00888 (9) | 0.0322 (5) | |
| C2 | 0.1832 (4) | 0.9866 (3) | 0.05590 (9) | 0.0342 (5) | |
| H2 | 0.0125 | 0.9702 | 0.0481 | 0.041* | |
| C3 | 0.2320 (7) | 1.1841 (3) | 0.07866 (13) | 0.0664 (9) | |
| H3A | 0.1459 | 1.2093 | 0.1199 | 0.100* | |
| H3B | 0.1820 | 1.2683 | 0.0436 | 0.100* | |
| H3C | 0.3990 | 1.1993 | 0.0868 | 0.100* | |
| C4 | 0.2478 (4) | 0.5021 (4) | 0.25784 (10) | 0.0344 (4) | |
| C5 | 0.0000 (3) | 0.5137 (3) | 0.22394 (9) | 0.0314 (5) |
| U11 | U22 | U33 | U12 | U13 | U23 | |
| N1 | 0.0392 (10) | 0.0462 (9) | 0.0223 (7) | −0.0056 (9) | 0.0030 (7) | −0.0009 (7) |
| O1 | 0.0435 (9) | 0.0828 (12) | 0.0269 (7) | 0.0179 (11) | 0.0048 (6) | 0.0122 (8) |
| O2 | 0.0390 (8) | 0.0535 (8) | 0.0308 (7) | 0.0133 (7) | 0.0067 (6) | 0.0025 (7) |
| O3 | 0.0209 (7) | 0.1291 (19) | 0.0369 (8) | 0.0020 (11) | 0.0013 (6) | 0.0228 (11) |
| O4 | 0.0315 (7) | 0.0865 (12) | 0.0286 (7) | 0.0002 (9) | −0.0048 (6) | 0.0114 (7) |
| O5 | 0.0320 (7) | 0.0665 (10) | 0.0246 (6) | −0.0041 (8) | −0.0033 (6) | 0.0080 (6) |
| O6 | 0.0203 (7) | 0.172 (2) | 0.0361 (8) | −0.0033 (13) | 0.0015 (6) | 0.0402 (12) |
| C1 | 0.0353 (10) | 0.0340 (9) | 0.0271 (9) | 0.0027 (8) | 0.0044 (8) | 0.0029 (8) |
| C2 | 0.0350 (10) | 0.0404 (10) | 0.0273 (9) | 0.0051 (11) | 0.0042 (8) | −0.0018 (8) |
| C3 | 0.105 (3) | 0.0392 (11) | 0.0552 (14) | 0.0083 (17) | 0.0140 (18) | −0.0081 (11) |
| C4 | 0.0212 (8) | 0.0548 (11) | 0.0273 (9) | −0.0004 (9) | −0.0006 (8) | 0.0063 (9) |
| C5 | 0.0222 (9) | 0.0469 (11) | 0.0252 (8) | −0.0011 (10) | −0.0007 (7) | 0.0060 (8) |
| N1—C2 | 1.483 (3) | O5—C5 | 1.219 (2) |
| N1—H1A | 0.8900 | O6—C5 | 1.235 (2) |
| N1—H1B | 0.8900 | C1—C2 | 1.508 (3) |
| N1—H1C | 0.8900 | C2—C3 | 1.522 (3) |
| O1—C1 | 1.303 (2) | C2—H2 | 0.9800 |
| O1—H1 | 0.8200 | C3—H3A | 0.9600 |
| O2—C1 | 1.205 (2) | C3—H3B | 0.9600 |
| O3—C4 | 1.297 (2) | C3—H3C | 0.9600 |
| O3—H3 | 0.8200 | C4—C5 | 1.548 (3) |
| O4—C4 | 1.199 (2) | ||
| C2—N1—H1A | 109.5 | C1—C2—H2 | 109.6 |
| C2—N1—H1B | 109.5 | C3—C2—H2 | 109.6 |
| H1A—N1—H1B | 109.5 | C2—C3—H3A | 109.5 |
| C2—N1—H1C | 109.5 | C2—C3—H3B | 109.5 |
| H1A—N1—H1C | 109.5 | H3A—C3—H3B | 109.5 |
| H1B—N1—H1C | 109.5 | C2—C3—H3C | 109.5 |
| C1—O1—H1 | 109.5 | H3A—C3—H3C | 109.5 |
| C4—O3—H3 | 109.5 | H3B—C3—H3C | 109.5 |
| O2—C1—O1 | 125.65 (18) | O4—C4—O3 | 126.7 (2) |
| O2—C1—C2 | 121.97 (18) | O4—C4—C5 | 122.01 (18) |
| O1—C1—C2 | 112.32 (17) | O3—C4—C5 | 111.32 (15) |
| N1—C2—C1 | 108.33 (16) | O5—C5—O6 | 126.65 (19) |
| N1—C2—C3 | 109.50 (18) | O5—C5—C4 | 118.05 (17) |
| C1—C2—C3 | 110.07 (19) | O6—C5—C4 | 115.29 (15) |
| N1—C2—H2 | 109.6 | ||
| O2—C1—C2—N1 | −29.3 (3) | O4—C4—C5—O5 | 180.0 (2) |
| O1—C1—C2—N1 | 153.29 (18) | O3—C4—C5—O5 | −1.0 (3) |
| O2—C1—C2—C3 | 90.4 (3) | O4—C4—C5—O6 | −1.0 (4) |
| O1—C1—C2—C3 | −87.0 (2) | O3—C4—C5—O6 | 178.1 (3) |
| D—H···A | D—H | H···A | D···A | D—H···A |
| N1—H1A···O6i | 0.89 | 1.91 | 2.728 (2) | 152 |
| N1—H1B···O4ii | 0.89 | 2.28 | 3.085 (2) | 150 |
| N1—H1C···O2iii | 0.89 | 2.30 | 2.978 (2) | 133 |
| N1—H1C···O5 | 0.89 | 2.35 | 2.896 (2) | 120 |
| O1—H1···O5iv | 0.82 | 1.76 | 2.575 (2) | 170 |
| O3—H3···O6v | 0.82 | 1.73 | 2.545 (2) | 172 |
| Symmetry codes: (i) −x, y+1/2, −z+1/2; (ii) −x+1, y+1/2, −z+1/2; (iii) x−1/2, −y+3/2, −z; (iv) x+1/2, −y+3/2, −z; (v) x+1, y, z. |
Experimental details
| Crystal data | |
| Chemical formula | C3H8NO2+·C2HO4− |
| Mr | 179.13 |
| Crystal system, space group | Orthorhombic, P212121 |
| Temperature (K) | 293 |
| a, b, c (Å) | 5.6304 (15), 7.2353 (15), 19.597 (3) |
| V (Å3) | 798.3 (2) |
| Z | 4 |
| Radiation type | Cu Kα |
| µ (mm−1) | 1.23 |
| Crystal size (mm) | 0.2 × 0.15 × 0.11 |
| Data collection | |
| Diffractometer | Enraf-Nonius sealed tube diffractometer |
| Absorption correction | ψ scan (North et al., 1968) |
| Tmin, Tmax | 0.79, 0.87 |
| No. of measured, independent and observed [I > 2σ(I)] reflections | 879, 879, 853 |
| Rint | 0.000 |
| (sin θ/λ)max (Å−1) | 0.601 |
| Refinement | |
| R[F2 > 2σ(F2)], wR(F2), S | 0.030, 0.083, 1.09 |
| No. of reflections | 879 |
| No. of parameters | 112 |
| H-atom treatment | H-atom parameters constrained |
| Δρmax, Δρmin (e Å−3) | 0.14, −0.14 |
| Absolute structure | See text |
Computer programs: CAD-4 Software (Enraf-Nonius, 1989), CAD-4 Software, SHELXS97 (Sheldrick, 1990), SHELXL97 (Sheldrick, 1997), PLATON (Spek, 1999), SHELXL97.
| N1—C2 | 1.483 (3) | O5—C5 | 1.219 (2) |
| O1—C1 | 1.303 (2) | O6—C5 | 1.235 (2) |
| O2—C1 | 1.205 (2) | C1—C2 | 1.508 (3) |
| O3—C4 | 1.297 (2) | C2—C3 | 1.522 (3) |
| O4—C4 | 1.199 (2) | C4—C5 | 1.548 (3) |
| O2—C1—C2—N1 | −29.3 (3) | O2—C1—C2—C3 | 90.4 (3) |
| O1—C1—C2—N1 | 153.29 (18) | O1—C1—C2—C3 | −87.0 (2) |
| D—H···A | D—H | H···A | D···A | D—H···A |
| N1—H1A···O6i | 0.89 | 1.91 | 2.728 (2) | 151.6 |
| N1—H1B···O4ii | 0.89 | 2.28 | 3.085 (2) | 150.4 |
| N1—H1C···O2iii | 0.89 | 2.30 | 2.978 (2) | 132.7 |
| N1—H1C···O5 | 0.89 | 2.35 | 2.896 (2) | 119.5 |
| O1—H1···O5iv | 0.82 | 1.76 | 2.575 (2) | 170.4 |
| O3—H3···O6v | 0.82 | 1.73 | 2.545 (2) | 171.6 |
| Symmetry codes: (i) −x, y+1/2, −z+1/2; (ii) −x+1, y+1/2, −z+1/2; (iii) x−1/2, −y+3/2, −z; (iv) x+1/2, −y+3/2, −z; (v) x+1, y, z. |




X-ray studies on crystalline complexes of amino acids with carboxylic acids have provided a wealth of information regarding intermolecular interactions and biomolecular aggregation patterns (Vijayan, 1988; Prasad & Vijayan, 1993). The crystal structures of glycinium oxalate (Subha Nandhini et al., 2001) and sarcosinium oxalate monohydrate (Krishnakumar et al., 1999) were elucidated in our laboratory. The present study reports the crystal structure of L-alaninium oxalate, (I), as part of a series of investigations being carried out to observe conformational changes in amino acid molecules and characteristic hydrogen-bonding patterns in their crystal structures.
Fig. 1 shows the molecular structure with the numering scheme. The alanine molecule exists in the cationic form with a positively charged amino group and an uncharged carboxylic acid group. The oxalic acid molecule exists in a mono-ionized state. The conformation of the L-alaninum cation about the N—Cα bond corresponds to the staggered ethane-type. A common feature among the crystal structures of glycinium oxalate and (I) is that the shortest cell dimensions are similar, 5.650 (2) and 5.6304 (15) Å, respectively. The semi-oxalate ions form hydrogen-bonded strings along the shortest cell axis, generated by translation, as in the structures of oxalic acid complexes of glycine (Subha Nandhini et al., 2001) and lysine (Venkatraman et al., 1997).
In the asymmetric unit, the L-alaninium cation and the semi-oxalate anion are linked to each other through a N—H···O hydrogen bond (Fig. 1). The head-to-tail hydrogen bond, with O2 of the carboxyl group as acceptor, observed among the amino acid molecules in the crystal structure may be described as a zigzag sequence along the 21 screw axis along the direction of the a axis. The alaninium and semi-oxalate ions form alternate columns leading to a layered arrangement parallel to the ac plane and each such layer is interconnected to the other through N—H···O hydrogen bonds. Two short C···O contacts involving the carboxyl oxygen of the alaninium ion [C1···O2(-1/2 + x, 3/2 - y, -z) = 2.931 (3) Å and C2···O2(-1/2 + x, 3/2 - y, -z) = 2.977 (3) Å] is also observed among these layers. The slight difference observed in the bond lengths of C5—O5 and C5—O6 in the carboxylate group of the semi-oxalate ion may be attributed to the difference in the strengths of the N—H···O hydrogen bonds in which both O5 and O6 are involved (Table 2). The overall aggregation pattern is distinctly different from that observed in the glycine–oxalic acid complex.